第7回「糖鎖科学統合データベース」
はじめに
我が国の糖鎖研究には長い歴史があり,現時点でも米国にひけをとっていない.多くのタンパク質は糖鎖修飾をうけ糖タンパク質として存在する.また,糖脂質やGPIアンカータンパク質(グリコシルホスファチジルイノシトールを結合し,これがアンカー役となり細胞膜につなぎとめられているタンパク質)も複合糖質であり,血液や細胞膜に存在するタンパク質のほとんどは糖タンパク質である[1].糖鎖修飾をうけてはじめてタンパク質は成熟し機能を獲得する[2-4].にもかかわらず,多くの研究者が糖鎖研究をさけてとおるのは,まだまだ基盤技術が一般化しておらず手を出しがたい研究分野だからであろう.遺伝子データベースやタンパク質データベースと比較して糖鎖に関するデータベースは未熟で内容にも乏しい.しかし,これまで,我が国がリードしてきたこの分野のデータベースを開発することは我が国の責務とも思える.糖鎖研究において初心者である研究者が解析の途中に糖鎖の壁に直面したとき,ユーザーフレンドリーなデータベースが存在したならどれだけ研究がやりやすくなるか.そのようなデータベースの構築をめざして,まずは,筆者らが過去10年間に産出してきた膨大な糖鎖研究のデータをまとめ公開することにした.さらに,国内に散在するデータを統合し,最終的には,アジア圏における糖鎖科学統合データベースを構築することを目的としている.
データベースの現状と問題点
最近,国家プロジェクトの成果を活用した糖鎖分析機器の普及がはじまったことから,研究者の利用できるデータの質が高くなってきた.筆者らの所属する産業技術総合研究所では,これまで,新エネルギー・産業技術総合開発機構(NEDO)の糖鎖合成関連遺伝子プロジェクト(GGプロジェクト)では,糖鎖関連遺伝子を網羅的に探索し,それぞれの糖転移酵素の基質特異性などの性状を解析してきた.糖鎖構造解析技術開発プロジェクト(SGプロジェクト)では,発見された糖鎖関連遺伝子のcDNAを活用し,組換え酵素を発現させて標準糖鎖の合成を行った.そして,この合成標準糖鎖とレクチンとの結合特異性を解析し,これまでにレクチンと100種類以上の糖鎖との相互作用情報を収集してきた.また,その標準糖鎖をイオントラップ式MALDI-TOF質量分析装置によるタンデム質量分析法(MSn法)により 質量分析装置を用いたMSnスペクトル計測することで,単糖の組成だけから構造を推定する方法とは異なり,グリコシド結合の様式を考慮して構造異性体(アイソバリック構造)を区別しながら糖鎖の構造を同定できるようになった.さらに,糖鎖機能解析技術開発プロジェクト(MGプロジェクト)では,これらの技術を組み合わせ,生理状態や病態の変動,治療に対する反応性などと相関する糖鎖関連バイオマーカーの探索を行い,有用なマーカーの実用化をめざしてきたという長い経緯がある.
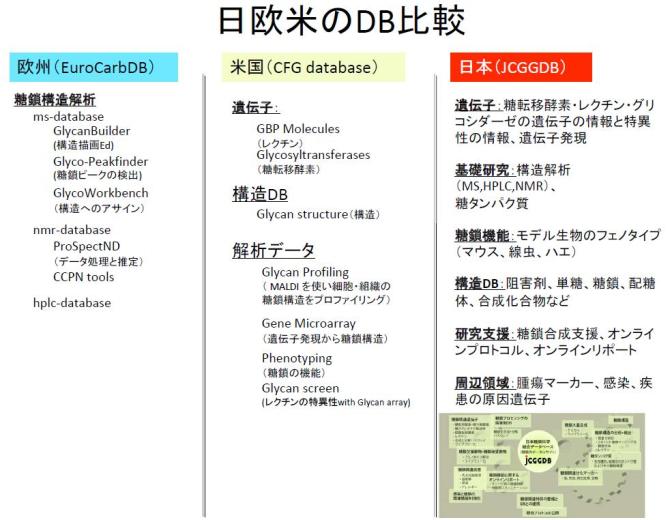
これらのNEDOプロジェクトと並行し,2007年度から2010年度まで文部科学省 統合データベースプロジェクトに参加することになった.このプロジェクトでは,国内の大学や研究機関などにある糖鎖に関連するデータベースを統合することが使命であった.そして,日本糖鎖科学コンソーシアム運営委員会,立命館大学,野口研究所,LipidBank構築委員会,生化学工業,名古屋大学,名古屋市立大学など,この研究領域の研究者の協力を得ながら,日本糖鎖科学コンソーシアム(Japan Consortium for Glycobiology and Glycotechnology:JCGG)のデータベースである日本糖鎖科学統合データベースJCGGDBを構築した.結果として,欧米のものと比較しても幅広い領域のデータベースを整備し公開することができたのである(図1).ここでは,さきに述べたNEDOプロジェクトの研究成果である糖鎖関連遺伝子とその基質特異性のデータベースGGDB [5],自動化フロンタルアフィニティークロマトグラフィーシステム[6] により得られたレクチンと糖鎖との相互作用のデータベースLfDB,標準糖鎖のMSnスペクトルデータベースGMDB,糖鎖付加位置特異的安定同位体標識法[7] により同定された糖タンパク質のデータベースGlycoProtDBを公開し,これらがJCGGDBの軸となった.この4つのデータベースは産業技術総合研究所が独自に開発した技術をもとに各種の試料を計測しデータベース化したものである.国内のデータベースの統合や連携を図りながら,実験データだけでは足りない情報を論文などから収集し,糖鎖と関連する分野のデータベースを増やしてきた.そして,それぞれのデータベースをキーワードや糖鎖構造により一括で検索できるよう横断検索機能を搭載した.糖鎖構造検索には米国に主導されるコンソーシアムCFG(Consortium for Functional Glycomics)の単糖シンボルによる構造描画機能や,化合物検索に適した化学分野のユーザーむきの描画機能を搭載した.また,単にデータベースを統合するだけではなく,これから糖鎖を研究したい人が糖鎖修飾された脂質やタンパク質を解析できるよう実験プロトコールの整備を行いGlycoPODとして公開した.
2011年度からは科学技術振興機構 統合データベースプロジェクト 統合化推進プログラムに参加することになった.まえのプロジェクトで整備したデータを利用し,協力機関である糖鎖生物学の知識ベースや抗糖鎖抗体の情報を多くもつ立命館大学,糖鎖の有機合成に強い野口研究所,糖鎖の立体化学に強い理化学研究所といっしょに,さらなる統合を推進していくことになった.これまでのデータの統合を進めると同時に,統合するための基盤整備と統合に必要なインフォマティクスのツールづくりに重点をおきながら,ウェット側の研究ニーズをみたせるよう目標をおいて構築を進めている.
ここでは,その現状をまとめることにする.海外では政府が我が国の予算を上回る金額を糖鎖の研究領域に積極的に投資している.とくに急成長を遂げているのは中国である.これとは対照的に,ヨーロッパではEuroCarb(European Carbohydrate)プロジェクトが数年前に終了し,我が国でも経済産業省による糖鎖に関連するNEDOプロジェクトが終了した.大型予算で残るのは科学技術振興機構ERATO伊藤グライコトリロジープロジェクトだけである.ヨーロッパの技術者や研究者は米国主導のコンソーシアムCFGに流れた.これまで先進的であった我が国の研究が先細り,数年後は糖鎖に関連する技術において優位な立場ではなくなる可能性がある.欧米にはEBI(European Bioinformatics Institute)や英国Imperial大学,米国Scripps研究所や米国Georgia大学Complex Carbohydrate Research Centerなどに糖鎖インフォマティクスの研究チームがあるが,我が国と同様にその人数は多くない.くわえて近い将来,糖鎖のデータを大量に産出するであろう中国や台湾にはゲノムやプロテオミクスのインフォマティストはいるが糖鎖のインフォマティストはいない.現状は,産業技術総合研究所がACGG(Asian Communications of Glycobiology and Glycotechnology)によるデータベースの構築を支援している.アジア圏における協力体制の強化のため,ACGGを主体とするデータベース連携の体制づくりをはじめている.
将来の理想像
糖鎖を研究するウェット研究者やバイオインフォマティスト,インフォマティシャンが少ないことから,糖鎖研究は技術的な面でいまだに発展途上であり,実験データを網羅しているとはいえない.少ないデータセットという状況においてウェット研究者が利用するという立場で考え,バイオインフォマティクスのツールやインタフェースの開発を進めていく必要がある.とくに開発しなければならないのは,つぎに示す内容である.統合化のためのデータやオントロジー(収集および標準化されたメタデータを記載し定義すること)の整備も必要ではあるが,実験データを理解するためのツール自体が不足しているので,実験データを理解したうえで新しいアルゴリズムを開発しなければ利用価値のあるプログラムは生まれないだろう.
- 糖鎖に関連したデータベースの統合化(標準化)
- 糖鎖の専門用語の整備および統一化
- 糖鎖のオントロジーの整備
- 糖鎖構造の整備(構造と名称の整備)
- 糖鎖科学の実験プロトコール
- 抗糖鎖抗体,レクチン,糖転移酵素,グリコシダーゼなどの糖鎖認識の理解
- 混合した糖ペプチドや糖タンパク質の質量分析による同定技術の開発
- 糖鎖の生合成経路および分解経路の解析と整備
- 糖鎖機能の情報集約
- 糖鎖関連疾患の情報集約
- 有機化学による糖鎖合成の情報集約
- 糖転移酵素やグリコシダーゼの阻害剤(漢方薬,配糖体を含む)の情報集約
- 微生物の糖転移酵素,グリコシダーゼ,レクチン
- 糖鎖関連特許情報の利用
おわりに
ゲノミクス→プロテオミクス→グライコミクス(プロテオグライコミクス)とオミックス研究が推移していくなか,糖鎖科学データベースに関してはぜひとも我が国から発信すべきであろう.もちろん最終的には,世界中のデータが統合された国際糖鎖科学統合データベースへと進化することを願っている.
参考文献
- Apweiler, R., Hermjakob, H. & Sharon, N.: On the frequency of protein glycosylation, as deduced from analysis of the SWISS-PROT database. Biochim. Biophys. Acta, 1473, 4-8 (1999) ↑
- Lee, T. K., Koh, A. S., Cui, Z. et al.: N-glycosylation controls functional activity of Oatp1, an organic anion transporter. Am. J. Physiol. Gastrointest. Liver Physiol., 285, G371-G381 (2003) ↑
- Asano, T., Katagiri, H., Takata, K. et al.: The role of N-glycosylation of GLUT1 for glucose transport activity. J. Biol. Chem., 266, 24632-24636 (1991) ↑
- Tsachaki, M., Serlidaki, D., Fetani, A. et al.: Glycosylation of BRI2 on asparagine 170 is involved in its trafficking to the cell surface but not in its processing by furin or ADAM10. Glycobiology, 21, 1382-1388 (2011) ↑
- Narimatsu, H.: Construction of a human glycogene library and comprehensive functional analysis. Glycoconj. J., 21, 17-24 (2004) ↑
- Tateno, H., Nakamura-Tsuruta, S. & Hirabayashi, J.: Frontal affinity chromatography: sugar-protein interactions. Nat. Protoc., 2, 2529-2537 (2007) ↑
- Kaji, H., Saito, H., Yamauchi, Y. et al.: Lectin affinity capture, isotope-coded tagging and mass spectrometry to identify N-linked glycoproteins. Nat. Biotechnol., 21, 667-672 (2003) ↑
↑ 押下で本文に戻ります。

Licensed under a Creative Commons 表示2.1日本 license ©2012 鹿内俊秀,成松久(産業技術総合研究所糖鎖医工学研究センター)
なお、本記事は細胞工学2012年2月号掲載の原稿を改変したものです。
